Ensemble NMA of E.coli DHFR structures
Background
Bio3D1 is an R package that provides interactive tools for structural bioinformatics. The primary focus of Bio3D is the analysis of bimolecular structure, sequence and simulation data (Grant et al. 2006).
Normal mode analysis (NMA) is one of the major simulation techniques used to probe large-scale motions in biomolecules. Typical application is for the prediction of functional motions in proteins. Version 2.0 of the Bio3D package now includes extensive NMA facilities (Skjaerven et al. 2015). These include a unique collection of multiple elastic network model force-fields, automated ensemble analysis methods, and variance weighted NMA (see also the NMA Vignette). Here we provide an in-depth demonstration of ensemble NMA with working code that comprise complete executable examples2.
Requirements
Detailed instructions for obtaining and installing the Bio3D package on various platforms can be found in the Installing Bio3D Vignette available both on-line and from within the Bio3D package. In addition to Bio3D the MUSCLE multiple sequence alignment program (available from the muscle home page must be installed on your system and in the search path for executables. Please see the installation vignette for further details.
About this document
This vignette was generated using Bio3D version 2.3.0.
Part I: Ensemble NMA of E.coli DHFR structures
In this vignette we perform ensemble NMA on the complete collection of E.coli Dihydrofolate reductase (DHFR) structures in the protein data-bank (PDB). Starting from only one PDB identifier (PDB ID 1rx2) we show how to search the PDB for related structures using BLAST, fetch and align the structures, and finally calculate the normal modes of each individual structure in order to probe for potential differences in structural flexibility.
Search and retrieve DHFR structures
Below we perform a blast search of the PDB database to identify related structures to our query E.coli DHFR sequence. In this particular example we use function get.seq() to fetch the query sequence for chain A of the PDB ID 1RX2 and use this as input to blast.pdb(). Note that get.seq() would also allow the corresponding UniProt identifier.
library(bio3d)
aa <- get.seq("1rx2_A")
blast <- blast.pdb(aa)
## Searching ... please wait (updates every 5 seconds) RID = YR4ZKKVY015
## .
## Reporting 576 hits
Function plot.blast() facilitates the visualization and filtering of the Blast results. It will attempt to set a seed position to the point of largest drop-off in normalized scores (i.e. the biggest jump in E-values). In this particular case we specify a cutoff (after initial plotting) of 225 to include only the relevant E.coli structures:
hits <- plot(blast, cutoff=225)
## * Possible cutoff values: 253 25
## Yielding Nhits: 125 576
##
## * Chosen cutoff value of: 225
## Yielding Nhits: 125
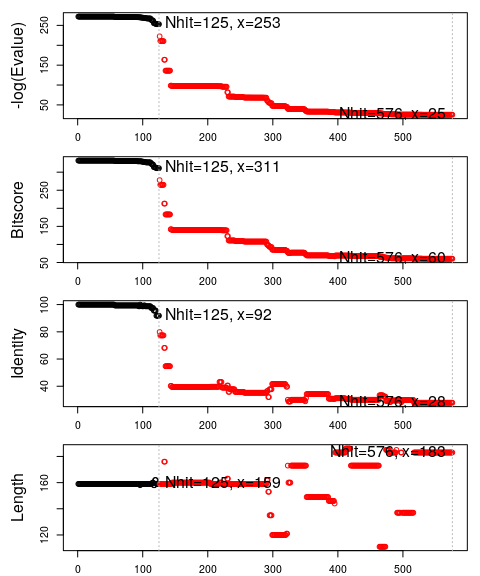
The Blast search and subsequent filtering identified a total of 101
related PDB structures to our query sequence. The PDB identifiers of
this collection are accessible through the pdb.id attribute to the
hits object (hits$pdb.id). Note that adjusting the cutoff argument
(to plot.blast()) will result in a decrease or increase of hits.
We can now use function get.pdb() and pdbslit() to fetch and parse the identified structures. Finally, we use pdbaln() to align the PDB structures.
# fetch PDBs
raw.files <- get.pdb(hits$pdb.id, path = "raw_pdbs", gzip=TRUE)
# split by chain ID
files <- pdbsplit(raw.files, ids = hits$pdb.id, path = "raw_pdbs/split_chain", ncore=4)
# align structures
pdbs.all <- pdbaln(files, fit=TRUE)
The pdbs.all object now contains aligned C-alpha atom data, including Cartesian coordinates, residue numbers, residue types, and B-factors. The sequence alignment is also stored by default to the FASTA format file 'aln.fa' (to view this you can use an alignment viewer such as SEAVIEW, see Requirements section above). In cases where manual edits of the alignment are necessary you can re-read the sequence alignment and coordinate data with:
aln <- read.fasta('aln.fa')
pdbs.all <- read.fasta.pdb(aln)
At this point the pdbs.all object contains all identified 101 structures. This might include structures with missing residues, and/or multiple structurally redundant conformers. For our subsequent NMA missing in-structure residues might bias the calculation, and redundant structures can be useful to omit to reduce the computational load. Below we inspect the connectivity of the PDB structures with a function call to inspect.connectivity(), and trim.pdbs() to filter out those structures from our pdbs.all object. Similarly, we omit structures that are conformationally redundant to reduce the computational load with function filter.rmsd():
# remove structures with missing residues
conn <- inspect.connectivity(pdbs.all, cut=4.05)
pdbs <- trim(pdbs.all, row.inds=which(conn))
# which structures are omitted
which(!conn)
# remove conformational redundant structures
rd <- filter.rmsd(pdbs$xyz, cutoff=0.1, fit=TRUE)
pdbs <- trim(pdbs, row.inds=rd$ind)
# remove "humanized" e-coli dhfr
excl <- unlist(lapply(c("3QL0", "4GH8"), grep, pdbs$id))
pdbs <- trim(pdbs, row.inds=which(!(1:length(pdbs$id) %in% excl)))
# a list of PDB codes of our final selection
ids <- unlist(strsplit(basename(pdbs$id), split=".pdb"))
Use print() to see a short summary of the pdbs object:
print(pdbs, alignment=FALSE)
##
## Call:
## trim.pdbs(pdbs = pdbs, row.inds = which(!(1:length(pdbs$id) %in%
## excl)))
##
## Class:
## pdbs, fasta
##
## Alignment dimensions:
## 108 sequence rows; 166 position columns (159 non-gap, 7 gap)
##
## + attr: id, xyz, resno, b, chain, ali, resid, call
Annotate collected PDB structures
Function pdb.annotate() provides a convenient way of annotating the PDB files we have collected. Below we use the function to annotate each structure to its source species. This will come in handy when annotating plots later on:
anno <- pdb.annotate(ids)
print(unique(anno$source))
## [1] "Escherichia coli" "Klebsiella pneumoniae"
Principal component analysis
A principal component analysis (PCA) can be performed on the structural ensemble (stored in the pdbs object) with function pca.xyz(). To obtain meaningful results we first superimpose all structures on the invariant core (function core.find()).
# find invariant core
core <- core.find(pdbs)
# superimpose all structures to core
pdbs$xyz = pdbfit(pdbs, core$c0.5A.xyz)
# identify gaps, and perform PCA
gaps.pos <- gap.inspect(pdbs$xyz)
gaps.res <- gap.inspect(pdbs$ali)
pc.xray <- pca.xyz(pdbs$xyz[,gaps.pos$f.inds])
# plot PCA
plot(pc.xray)
Note that a call to the wrapper function pca() would provide identical results as the above code:
pc.xray <- pca(pdbs, core.find=TRUE)
Function rmsd() will calculate all pairwise RMSD values of the structural ensemble. This facilitates clustering analysis based on the pairwise structural deviation:
rd <- rmsd(pdbs)
hc.rd <- hclust(dist(rd))
grps.rd <- cutree(hc.rd, k=4)
plot(pc.xray$z[,1:2], col="grey50", pch=16, cex=1.3,
ylab="Prinipcal Component 2", xlab="Principal Component 1")
points(pc.xray$z[,1:2], col=grps.rd, pch=16, cex=0.9)

To visualize the major structural variations in the ensemble function mktrj() can be used to generate a trajectory PDB file by interpolating along the eigenvector:
mktrj(pc.xray, pc=1,
resno=pdbs$resno[1, gaps.res$f.inds],
resid=pdbs$resid[1, gaps.res$f.inds])
Function pdbfit() can be used to write the PDB files to separate directories according to their cluster membership:
pdbfit(trim(pdbs, row.inds=which(grps.rd==1)), outpath="grps1") ## closed
pdbfit(trim(pdbs, row.inds=which(grps.rd==2)), outpath="grps2") ## open
pdbfit(trim(pdbs, row.inds=which(grps.rd==3)), outpath="grps3") ## occluded
Normal modes analysis
Function nma.pdbs() will calculate the normal modes of each protein
structure stored in the pdbs object. The normal modes are calculated
on the full structures as provided by object pdbs. With the default
argument rm.gaps=TRUE unaligned atoms are omitted from output:
modes <- nma(pdbs, rm.gaps=TRUE, ncore=4)
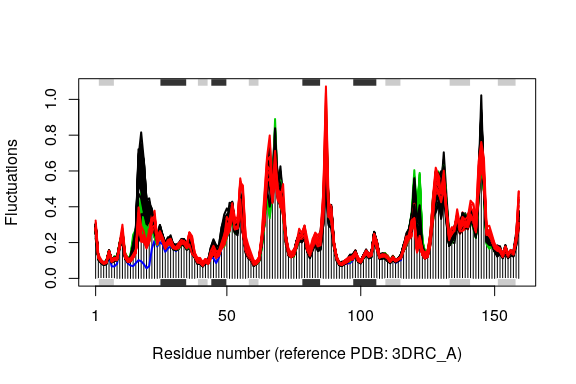
The modes object of class enma contains aligned normal mode data including fluctuations, RMSIP data, and aligned eigenvectors. A short summary of the modes object can be obtain by calling the function print(), and the aligned fluctuations can be plotted with function plot.enma().
print(modes)
##
## Call:
## nma.pdbs(pdbs = pdbs, rm.gaps = TRUE, ncore = 4)
##
## Class:
## enma
##
## Number of structures:
## 108
##
## Attributes stored:
## - Root mean square inner product (RMSIP)
## - Aligned atomic fluctuations
## - Aligned eigenvectors (gaps removed)
## - Dimensions of x$U.subspace: 477x471x108
##
## Coordinates were aligned prior to NMA calculations
##
## + attr: fluctuations, rmsip, U.subspace, L, full.nma, xyz,
## call
# plot modes fluctuations
plot(modes, pdbs=pdbs, col=grps.rd, label=NULL)
## Re-reading PDB (3DRC_A) to extract SSE
## Warning in dssp.pdb(pdb.ref, ...): Non-protein residues detected in input
## PDB: CL, MTX, HOH
hc.nma <- hclust(as.dist(1-modes$rmsip))
grps.nma <- cutree(hc.nma, k=4)
heatmap(1-modes$rmsip, distfun = as.dist, labRow = ids, labCol = ids,
ColSideColors=as.character(grps.nma), RowSideColors=as.character(grps.nma))

Fluctuation analysis
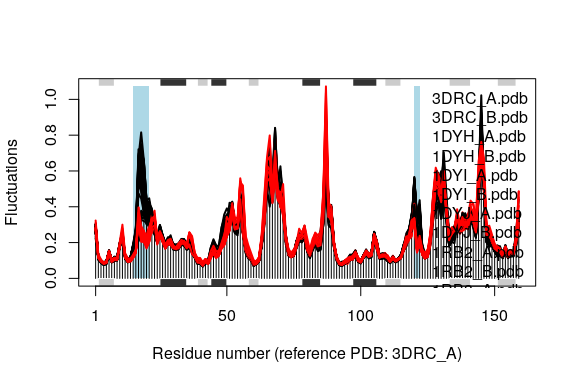
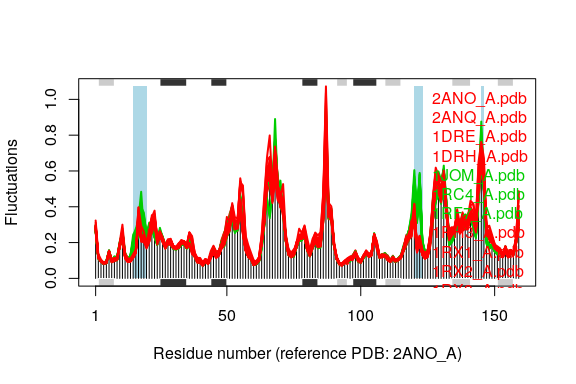
Comparing the mode fluctuations of two groups of structures can reveal specific regions of distinct flexibility patterns. Below we focus on the differences between the open (black), closed (red) and occluded (green) conformations of the E.coli structures:
cols <- grps.rd
cols[which(cols!=1 & cols!=2)]=NA
plot(modes, pdbs=pdbs, col=cols, signif=TRUE)
## Re-reading PDB (3DRC_A) to extract SSE
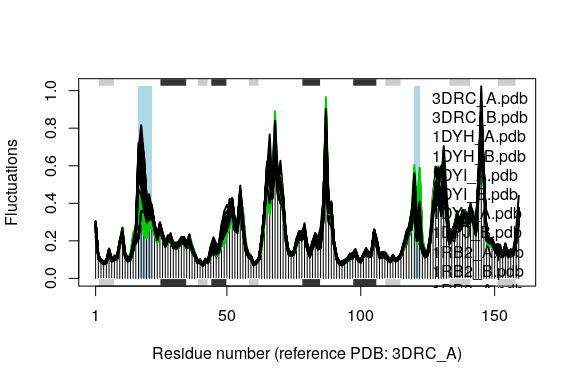
cols <- grps.rd
cols[which(cols!=1 & cols!=3)]=NA
plot(modes, pdbs=pdbs, col=cols, signif=TRUE)
## Re-reading PDB (3DRC_A) to extract SSE
cols <- grps.rd
cols[which(grps.rd!=2 & grps.rd!=3)]=NA
plot(modes, pdbs=pdbs, col=cols, signif=TRUE)
## Re-reading PDB (2ANO_A) to extract SSE
## PDB has ALT records, taking A only, rm.alt=TRUE
Compare with MD simulation
The above analysis can also nicely be integrated with molecular dynamics (MD) simulations. Below we read in a 5 ns long MD trajectory (of PDB ID 1RX2). We visualize the conformational sampling by projecting the MD conformers onto the principal components (PCs) of the X-ray ensemble, and finally compare the PCs of the MD simulation to the normal modes:
pdb <- read.pdb("md-traj/1rx2-CA.pdb")
trj <- read.ncdf("md-traj/1rx2_5ns.nc")
inds <- pdb2aln.ind(pdbs, pdb, gaps.res$f.inds)
inds.core <- pdb2aln.ind(pdbs, pdb, core$c0.5A.atom)
# inds$a is for the pdbs object
# inds$b is for the MD structure
trj.fit <- fit.xyz(pdbs$xyz[1,], trj, core$c0.5A.xyz, inds.core$b$xyz)
proj <- project.pca(trj.fit[, inds$b$xyz], pc.xray)
cols <- densCols(proj[,1:2])
plot(proj[,1:2], col=cols, pch=16,
ylab="Prinipcal Component 2", xlab="Principal Component 1",
xlim=range(pc.xray$z[,1]), ylim=range(pc.xray$z[,2]))
points(pc.xray$z[,1:2], col=1, pch=1, cex=1.1)
points(pc.xray$z[,1:2], col=grps.rd, pch=16)
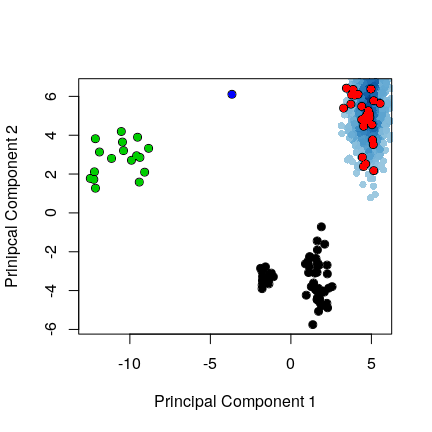
# PCA of the MD trajectory
pc.md <- pca.xyz(trj.fit[, inds$b$xyz])
# compare MD-PCA and NMA
r <- rmsip(pc.md$U, modes$U.subspace[,,1])
print(r)
## $overlap
## b1 b2 b3 b4 b5 b6 b7 b8 b9 b10
## a1 0.529 0.008 0.003 0.027 0.000 0.000 0.001 0.001 0.000 0.019
## a2 0.017 0.070 0.019 0.005 0.026 0.027 0.006 0.000 0.014 0.057
## a3 0.035 0.207 0.006 0.004 0.097 0.002 0.006 0.010 0.001 0.033
## a4 0.003 0.165 0.139 0.105 0.045 0.018 0.051 0.052 0.009 0.000
## a5 0.003 0.013 0.011 0.008 0.065 0.003 0.009 0.049 0.008 0.001
## a6 0.002 0.046 0.000 0.031 0.041 0.082 0.018 0.082 0.059 0.000
## a7 0.003 0.086 0.106 0.071 0.004 0.001 0.047 0.009 0.051 0.003
## a8 0.018 0.023 0.220 0.331 0.027 0.102 0.006 0.003 0.000 0.005
## a9 0.082 0.083 0.007 0.000 0.009 0.001 0.064 0.069 0.011 0.001
## a10 0.001 0.003 0.121 0.011 0.193 0.006 0.015 0.002 0.002 0.005
##
## $rmsip
## [1] 0.6443585
##
## attr(,"class")
## [1] "rmsip"
plot(r, xlab="MD PCA", ylab="NMA")
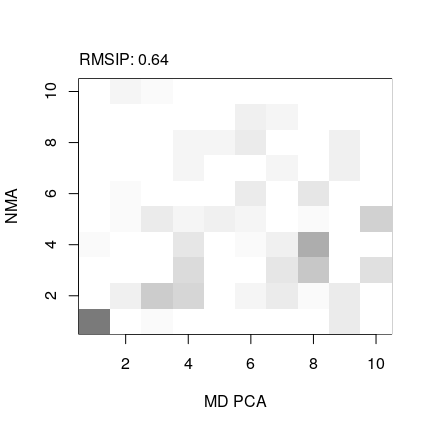
# compare MD-PCA and X-rayPCA
r <- rmsip(pc.md, pc.xray)
Domain analysis with GeoStaS
Identification of regions in the protein that move as rigid bodies is
facilitated with the implementation of the GeoStaS algorithm
(Romanowska, Nowinski, and Trylska 2012). Below we demonstrate the use
of function geostas() on data obtained from ensemble NMA, an
ensemble of PDB structures, and a 5 ns long MD simulation. See
help(geostas) for more details and further examples.
GeoStaS on a PDB ensemble: Below we input the pdbs object to
function geostas() to identify dynamic domains. Here, we attempt to
divide the structure into 2 sub-domains using argument k=2. Function
geostas() will return a grps attribute which corresponds to the
domain assignment for each C-alpha atom in the structure. Note that we
use argument fit=FALSE to avoid re-fitting the coordinates since.
Recall that the coordinates of the pdbs object has already been
superimposed to the identified invariant core (see above). To visualize
the domain assignment we write a PDB trajectory of the first principal
component (of the Cartesian coordinates of the pdbs object), and add
argument chain=gs.xray$grps to replace the chain identifiers with the
domain assignment:
# Identify dynamic domains
gs.xray <- geostas(pdbs, k=2, fit=FALSE)
# Visualize PCs with colored domains (chain ID)
mktrj(pc.xray, pc=1, chain=gs.xray$grps)
GoeStaS on ensemble NMA: We can also identify dynamic domains from
the normal modes of the ensemble of 82 protein structures stored in the
modes object. By using function mktrj.enma() we generate a
trajectory from the first five modes for all 82 structures. We then
input this trajectory to function geostas():
# Build conformational ensemble
trj.nma <- mktrj.enma(modes, pdbs, m.inds=1:5, s.inds=NULL, mag=10, step=2, rock=FALSE)
trj.nma
##
## Total Frames#: 5940
## Total XYZs#: 477, (Atoms#: 159)
##
## [1] 25.125 59.135 5.302 <...> 33.862 47.305 6.474 [2833380]
##
## + attr: Matrix DIM = 5940 x 477
# Fit to invariant core
trj.nma <- fit.xyz(trj.nma[1,], trj.nma,
fixed.inds=core$c0.5A.xyz,
mobile.inds=core$c0.5A.xyz)
# Run geostas to find domains
gs.nma <- geostas(trj.nma, k=2, fit=FALSE)
## .. 'xyz' coordinate data with 5940 frames
## .. coordinates are not superimposed prior to geostas calculation
## .. calculating atomic movement similarity matrix ('amsm.xyz()')
## .. dimensions of AMSM are 159x159
## .. clustering AMSM using 'kmeans'
# Write NMA generated trajectory with domain assignment
write.pdb(xyz=trj.nma, chain=gs.nma$grps)
Note that function geostas.enma() wraps these three main functions
calls to faciliatet this calculation. See help(geostas) for more
information.
GeoStaS on a MD trajectory: The domain analysis can also be
performed on the trajectory data obtained from the MD simulation (see
above). Recall that the MD trajectory has already been superimposed to
the invariant core. We therefore use argument fit=FALSE below. We then
perform a new PCA of the MD trajectory, and visualize the domain
assingments with function mktrj():
gs.md <- geostas(trj, k=2, fit=FALSE)
pc.md <- pca(trj, fit=FALSE)
mktrj(pc.md, pc=1, chain=gs.md$grps)
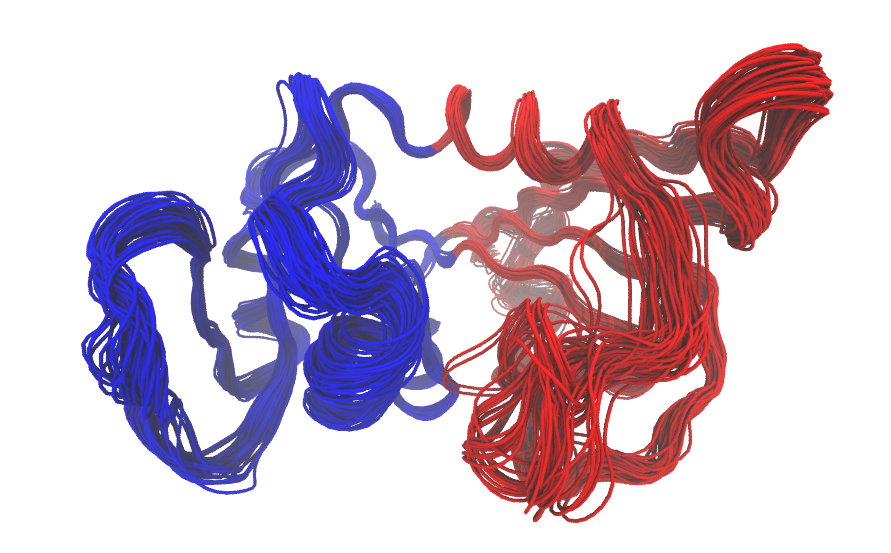
Measures for modes comparison
Bio3D now includes multiple measures for the assessment of similarity between two normal mode objects. This enables clustering of related proteins based on the predicted modes of motion. Below we demonstrate the use of root mean squared inner product (RMSIP), squared inner product (SIP), covariance overlap, bhattacharyya coefficient, and PCA of the corresponding covariance matrices.
# Similarity of atomic fluctuations
sip <- sip(modes)
hc.sip <- hclust(as.dist(1-sip), method="ward.D2")
grps.sip <- cutree(hc.sip, k=3)
hclustplot(hc.sip, k=3, colors=grps.rd, labels=ids, cex=0.7, main="SIP", fillbox=FALSE)
par(fig=c(.65, 1, .45, 1), new = TRUE)
plot(pc.xray$z[,1:2], col="grey50", pch=16, cex=1.1,
ylab="", xlab="", axes=FALSE, bg="red")
points(pc.xray$z[,1:2], col=grps.sip, pch=16, cex=0.7)
box()
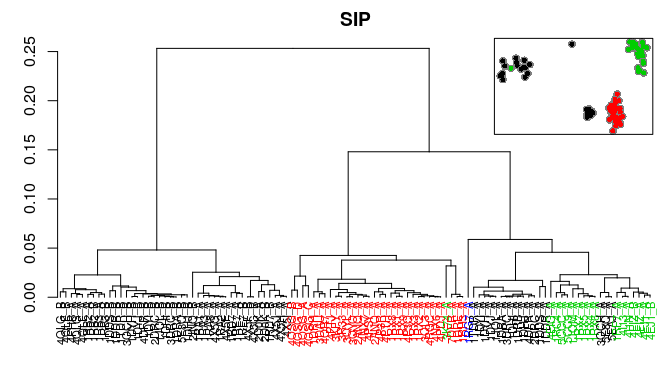
# RMSIP
rmsip <- rmsip(modes)
hc.rmsip <- hclust(dist(1-rmsip), method="ward.D2")
grps.rmsip <- cutree(hc.rmsip, k=3)
hclustplot(hc.rmsip, k=3, colors=grps.rd, labels=ids, cex=0.7, main="RMSIP", fillbox=FALSE)
par(fig=c(.65, 1, .45, 1), new = TRUE)
plot(pc.xray$z[,1:2], col="grey50", pch=16, cex=1.1,
ylab="", xlab="", axes=FALSE)
points(pc.xray$z[,1:2], col=grps.rmsip, pch=16, cex=0.7)
box()
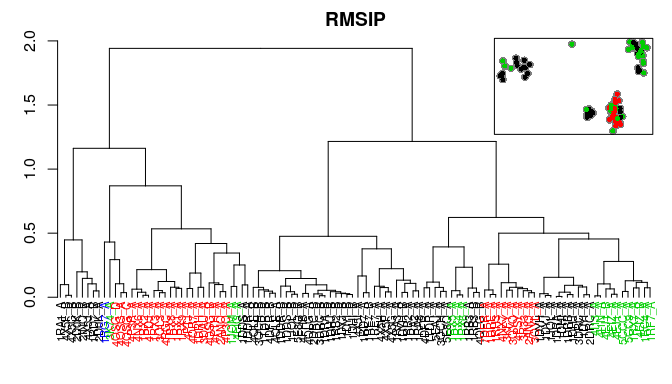
# Covariance overlap
co <- covsoverlap(modes, subset=200)
hc.co <- hclust(as.dist(1-co), method="ward.D2")
grps.co <- cutree(hc.co, k=3)
hclustplot(hc.co, k=3, colors=grps.rd, labels=ids, cex=0.7, main="Covariance overlap", fillbox=FALSE)
par(fig=c(.65, 1, .45, 1), new = TRUE)
plot(pc.xray$z[,1:2], col="grey50", pch=16, cex=1.1,
ylab="", xlab="", axes=FALSE)
points(pc.xray$z[,1:2], col=grps.co, pch=16, cex=0.7)
box()
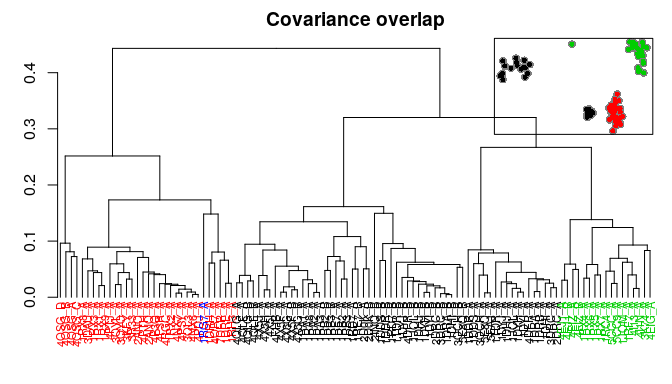
# Bhattacharyya coefficient
covs <- cov.enma(modes)
bc <- bhattacharyya(modes, covs=covs)
hc.bc <- hclust(dist(1-bc), method="ward.D2")
grps.bc <- cutree(hc.bc, k=3)
hclustplot(hc.bc, k=3, colors=grps.rd, labels=ids, cex=0.7, main="Bhattacharyya coefficient", fillbox=FALSE)
par(fig=c(.65, 1, .45, 1), new = TRUE)
plot(pc.xray$z[,1:2], col="grey50", pch=16, cex=1.1,
ylab="", xlab="", axes=FALSE)
points(pc.xray$z[,1:2], col=grps.bc, pch=16, cex=0.7)
box()
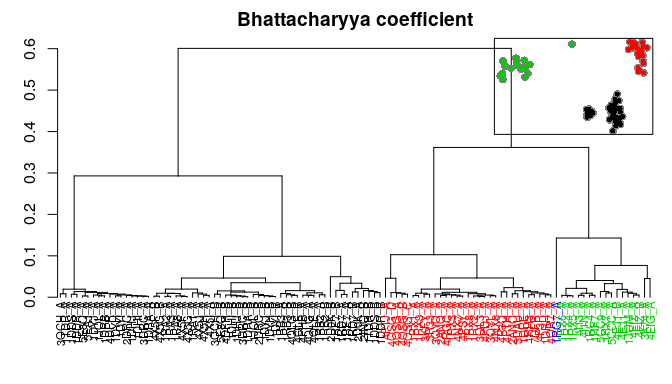
# PCA of covariance matrices
pc.covs <- pca.array(covs)
hc.covs <- hclust(dist(pc.covs$z[,1:2]), method="ward.D2")
grps.covs <- cutree(hc.covs, k=3)
hclustplot(hc.covs, k=3, colors=grps.rd, labels=ids, cex=0.7, main="PCA of covariance matrices", fillbox=FALSE)
par(fig=c(.65, 1, .45, 1), new = TRUE)
plot(pc.xray$z[,1:2], col="grey50", pch=16, cex=1.1,
ylab="", xlab="", axes=FALSE)
points(pc.xray$z[,1:2], col=grps.covs, pch=16, cex=0.7)
box()

Document Details
This document is shipped with the Bio3D package in both R and PDF formats. All code can be extracted and automatically executed to generate Figures and/or the PDF with the following commands:
library(rmarkdown)
render("Bio3D_nma-dhfr-partI.Rmd", "all")
Information About the Current Bio3D Session
print(sessionInfo(), FALSE)
## R version 3.3.1 (2016-06-21)
## Platform: x86_64-redhat-linux-gnu (64-bit)
## Running under: Fedora 24 (Twenty Four)
##
## attached base packages:
## [1] stats graphics grDevices utils datasets methods base
##
## other attached packages:
## [1] rmarkdown_1.0 bio3d_2.3-0
##
## loaded via a namespace (and not attached):
## [1] Rcpp_0.12.7 codetools_0.2-14 digest_0.6.10
## [4] grid_3.3.1 formatR_1.4 magrittr_1.5
## [7] evaluate_0.9 ncdf4_1.15 KernSmooth_2.23-15
## [10] stringi_1.1.1 tools_3.3.1 stringr_1.0.0
## [13] yaml_2.1.13 parallel_3.3.1 htmltools_0.3.5
## [16] knitr_1.14
References
Grant, B.J., A.P.D.C Rodrigues, K.M. Elsawy, A.J. Mccammon, and L.S.D. Caves. 2006. “Bio3d: An R Package for the Comparative Analysis of Protein Structures.” Bioinformatics 22: 2695–6. doi:10.1093/bioinformatics/btl461.
Romanowska, Julia, Krzysztof S. Nowinski, and Joanna Trylska. 2012. “Determining geometrically stable domains in molecular conformation sets.” Journal of Chemical Theory and Computation 8 (8): 2588–99. doi:10.1021/ct300206j.
Skjaerven, L., X.Q. Yao, G. Scarabelli, and B.J. Grant. 2015. “Integrating Protein Structural Dynamics and Evolutionary Analysis with Bio3D.” BMC Bioinformatics 15: 399. doi:10.1186/s12859-014-0399-6.
-
The latest version of the package, full documentation and further vignettes (including detailed installation instructions) can be obtained from the main Bio3D website: http://thegrantlab.org/bio3d/ ↩
-
This vignette contains executable examples, see
help(vignette)for further details. ↩